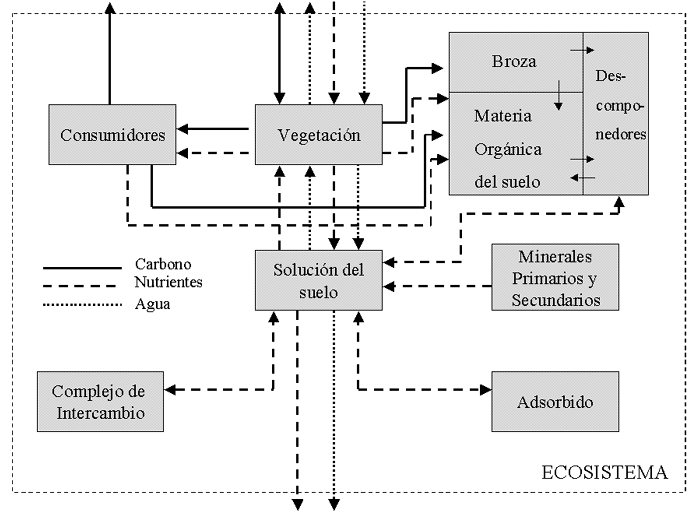
Descripción general:
El picaflor de Arica (
Eulidia yarrellii) es la especie de ave más pequeña del país y uno de los picaflores de menor
tamaño en el mundo. Pertenece al Orden
Apodiforme, Familia
Trochilidae y Subfamilia
Trochilinae. Presenta una longitud de 7 a 8 cm en hembras y 8 a 9 centímetros en machos, y su peso no sobrepasa los 4 gramos. (DPRNR, 2004). Su
alimentación consiste en néctar e insectos que caza en vuelo y construye nidos donde deposita dos huevos en
promedio. Según Aves de Chile, esta especie posee un notorio dimorfismo sexual:
Macho: Cabeza y partes superiores verdes oliva metálicos, garganta iridiscente, rojo púrpura vista de costado y negro vista de frente; resto de las partes inferiores blanco acanelado. Cola con rectrices centrales verde oliva metálico; las externas delgadas con color café negruzco.
(Ver imagen 1)
Hembra: Garganta sin rojo púrpura; con toda la parte inferior blanco acanelado, haciéndose más café hacia los flancos y subcaudales. Rectrices centrales verde metálicos, y las externas con barba externa café y punta blanca.
(Ver imagen 2)
Imagen 1: Picaflor de Arica Macho
Fuente: Cañas J. ,Aves de Chile, 2015.
Imagen 2: Picaflor de Arica Hembra
Fuente: Ávila M. ,Aves de Chile, 2015.
Este picaflor es endémico de los valles del desierto del norte de Chile y del sur del Perú. Su hábitat está bastante relacionado con los oasis
vegetales, valles cercanos y ocasionalmente las zonas costeras, siendo relevante
la presencia de árboles y flores productoras de néctar, dada su particular
alimentación. (DPRNR, 2014)
Registros antiguos de la especie indican que si bien poseía una distribución restringida, localmente
era muy abundante. Sin embargo, a partir de la década de los ‘70, la frecuencia en los registros disminuyó,
llegando a niveles considerados extremadamente bajos en los últimos años. Actualmente, se encuentra catalogado como
En Peligro (IUCN, 2014).
Posibles causas del decline de la población:
Según Aves de Chile, esta especie fue descrita como muy abundante a principios del siglo pasado, aunque en el año 2003 se estimó una población de 1500 individuos, pasando luego a conformar una población de menos de 500 individuos en el censo del 2012, por lo que fue catalogada como
En peligro. Frente a este hecho, una posible causa de la disminución progresiva es la utilización de los sectores pertenecientes al hábitat de la especie para la producción agrícola, sectorizando los lugares disponibles para realizar su ciclo de vida de manera natural, lo que finalmente desencadena una migración a otras zonas, las cuales no necesariamente presentan las características necesarias para que la población siga aumentando en número, principalmente por su reducida alimentación, aunque este picaflor posee la capacidad para alimentarse de otras
especies de carácter exótico, como la
lantana o cultivos agrícolas como
alfalfa o tomate (Seremi de Medio Ambiente, 2013).
Por otro lado, la introducción del Picaflor de Cora (
Thaumastura cora) en su hábitat, genera una posible competencia por los recursos del área, principalmente el alimento, debido a que lo supera en tamaño (mide aproximadamente de 12 a 13 cms) y número (es catalogado como
muy abundante, principalmente en el sector peruano en el cual también habita el Picaflor de Arica).
Tabla 1: Características, niveles jerárquicos y función del Picaflor de Arica.
Atributo
|
Nivel
jerárquico
|
Tipo
|
Peso
|
Individuo
|
Composicional
|
Tamaño poblacional
|
Población
|
Composicional
|
Distribución
|
Población
|
Estructural
|
Depredación/competencia
|
Comunidad
|
Funcional
|
Reproducción
|
Individuo
|
Funcional
|
Población mínima viable
|
Población
|
Funcional
|
Productividad de néctar
|
Población
|
Funcional
|
Fuente: Elaboración propia, 2015.
El Picaflor de Arica y el proceso de evolución por selección natural:
Este picaflor inicialmente presentó variadas características adaptativas al medio en el cual se desenvolvía. Si bien este actualmente se encuentra marginado, por las posibles causas que están generando un decline en la población de esta especie, estas aves se encuentran en un posible peligro de extinción, no tan sólo por la reducción de su hogar por efecto antrópico, sino también por la introducción del Picaflor de Cora, debido a que posee "mayor flexibilidad y perseverancia para captar los recursos lo hacen más apto para las condiciones ambientales en los cuales esta inmerso", según declara el Seremi de Medio Ambiente en su texto
Picaflor de Arica.
Los dos factores anteriormente mencionados, posicionan a esta especie a un nuevo proceso de adaptación o extinción, donde los individuos con mejor adaptación a un nuevo ambiente o que posean las características para establecer una competencia efectiva con la otra especie residente en el mismo entorno sobrevivirán y perpetuarán estos genes a las generaciones siguientes. Si no se logran reproducir estos individuos a una tasa más o menos elevada (por la poca densidad poblacional existente actualmente), la especie no logrará sobrevivir a las condiciones en las cuales vive ahora, llevándose a cabo el proceso de selección natural basado en la evolución.
Bibliografía:
Departamento de protección de los Recursos naturales
renovables (2004) Programa para la Recuperación del Picaflor de Arica e
inventario de su tamaño poblacional. DEPROREN. SAG. URL:
http://boletindeproren.sag.gob.cl/julio2004/archivos_pdf/picaflor.pdf (acceso
Octubre 03 2015)
Herreros de Lartundo, J. , Ch. Tala González y C. Estades
Marfán. (ed). 2013. Picaflor de Arica, el ave más amenazada de Chile.
Ministerio del Medio Ambiente. Arica, Chile. 84 pp.